































DNR ir RNR molekulinńó struktŇęra
- 1. DNR IR RNRDNR IR RNR MOLEKULINńĖMOLEKULINńĖ STRUKTŇ™RASTRUKTŇ™RA
- 2. ńģVADAS ÔĀģ Genetikos kursńÖ pradńósime nuo molekulinńós genetikos ÔĀģ Molekulinńó genetika tyrinńója DNR struktŇęrńÖ ir funkcijas molekuliŇ≥ lygmenyje ÔĀģ PastarŇ≥jŇ≥ metŇ≥ mokslo pasiekimai, tobulinant tyrimo metodus ir kuriant naujas koncepcijas, labai iŇ°plńótńó molekulinńós genetikos Ňĺinias ÔĀģ ҆is progresas palietńó ne tik molekulinńô, tańćĺĪ≤Ļu ir poŇĺymiŇ≥ perdavimo bei populiacijŇ≥ genetikas ÔĀģ DidŇĺiaja dalimi genetikos Ňĺinios yra sukauptos tiriant DNR ir RNR 2-2
- 3. ÔĀģ Savo funkcijńÖ genetinńó medŇĺiaga atlieka tik tada, kai ji pasiŇĺymi tam tikromis savybńómis ÔĀģ 1. Informacija: Ji privalo turńóti informacijńÖ, bŇętinńÖ visam organizmui sukurti ÔĀģ 2. Paveldńójimu: Ji privalo bŇęti perduodama iŇ° tńóvŇ≥ vaikams ÔĀģ 3. Replikacija: Ji turi bŇęti kopijuojama ÔĀģ Tada ji gali bŇęti perduodama iŇ° tńóvŇ≥ vaikams ÔĀģ 4. Kintamumu: Ji privalo gebńóti keistis ÔĀģ Toks kintamumas uŇĺtikrina plańćĺĪ≤Ļi stebima fenotpinńô variacijńÖ kiekvienos rŇęŇ°ies viduje 2-3 2.1 DNR KAIP GENETINńĖ MEDŇĹIAGA
- 4. ÔĀģ Daugelio genetikŇ≥, tarp jŇ≥ ir Mendelio, duomenys atitiko Ň°ias keturias savybes ÔĀģ TańćĺĪ≤Ļu genetinńós medŇĺiagos cheminńó prigimtis negalńójo bŇęti nustatyta vien tik kryŇĺminant organizmus ÔĀģ IŇ° tikrŇ≥jŇ≥, DNR kaip genetinńós medŇĺiagos identifikavimas pareikalavo naujoviŇ°ko poŇĺiŇęrio ńĮ eksperimentinńô biologijńÖ 2-4 2.1 DNR KAIP GENETINńĖ MEDŇĹIAGA
- 5. ÔĀģ Griffithas tyrńó bakterijńÖ (Diplococcus pneumoniae), kuri dabar vadinama Streptococcus pneumoniae ÔĀģ Yra aptinkami du S. pneumoniae kamienai ÔĀģ S ÔɆ Smooth (lygus) ÔĀģ Sekretuoja polisacharidinńô kapsulńô ÔĀģ ҆i apsaugo bakterijńÖ nuo gyvŇęnŇ≥ imuninńós sistemos ÔĀģ Auginamos ant kietos terpńós suformuoja lygias kolonijas ÔĀģ R ÔɆ Rough (Ň°iurkŇ°tus) ÔĀģ Nesugeba sekretuoti kapsulńós ÔĀģ KolonijŇ≥ kraŇ°tai nelygŇęs 2-5 Fredericko Griffitho eksperimentai su Streptococcus pneumoniae
- 6. ÔĀģ Be to, dviejŇ≥ S kamienŇ≥ kapsulńós gali reikŇ°mingai skirtis pagal savo cheminńô sudńótńĮ 2-6 ÔĀģ Retos mutacijos gali paversti S kamienus R kamienais ir atvirkŇ°ńćĺĪ≤Ļi ÔĀģ TańćĺĪ≤Ļu mutacjos nekeińćĺĪ≤Ļ kamienŇ≥ tipŇ≥
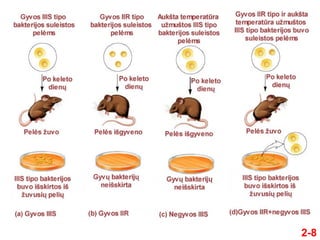
- 7. 2-7 ÔĀģ 1928 m. Griffithas atliko eksperimentus naudodamas du S. pneumoniae kamienus: IIIS tipo ir IIR tipo ÔĀģ 1. Pelńóms suleistos gyvos IIIS tipo bakterijos ÔĀģ Pelńós Ňĺuvo ÔĀģ IIIS tipo bakterijos buvo iŇ°skirtos iŇ° peliŇ≥ kraujo ÔĀģ 2. Pelńóms suleistos gyvos IIR tipo bakterijos ÔĀģ Pelńós iŇ°gyveno ÔĀģ IŇ° peliŇ≥ kraujo nebuvo iŇ°skirta gyvŇ≥ bakterijŇ≥ ÔĀģ 3. Pelńóms suleistos aukŇ°ta temperatŇęra uŇĺmuŇ°tos IIIS tipo bakterijos ÔĀģ Pelńós iŇ°gyveno ÔĀģ IŇ° peliŇ≥ kraujo nebuvo iŇ°skirta gyvŇ≥ bakterijŇ≥ ÔĀģ 4. Pelńóms suleistos gyvos IIR tipo aukŇ°ta temperatŇęra uŇĺmuŇ°tos IIIS tipo bakterijos ÔĀģ Pelńós Ňĺuvo ÔĀģ IIIS tipo bakterijos buvo iŇ°skirtos iŇ° peliŇ≥ kraujo
- 8. 2-8
- 9. 2-9 ÔĀģ Griffithas nusprendńó, kad kaŇĺkokia medŇĺiaga iŇ° ŇĺuvusiŇ≥ IIIS tipo bakterijŇ≥ transformavo IIR tipo bakterijas ńĮ IIIS tipńÖ ÔĀģ Jis pavadino Ň°ńĮ procesńÖ transformacija ÔĀģ MedŇĺiaga, kuri tai padarńó, buvo pavadinta transformuojanńćińÖja prieŇĺastimi ÔĀģ Griffithui nepavyko nustatyti, kokia tai medŇĺiaga ÔĀģ Transformuojanńćiosios prieŇĺasties prigimtis buvo nustatyta gerokai vńóliau, naudojant ńĮvairius biocheminius metodus
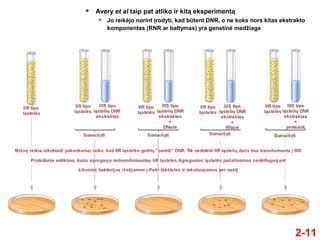
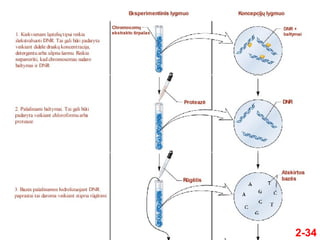
- 10. ÔĀģ Avery, MacLeod ir McCarty suprato, kad Griffitho tyrimŇ≥ rezultatai gali bŇęti panaudoti genetinei medŇĺiagai nustatyti ÔĀģ Jie atliko savo eksperimentus penktame XX a. deŇ°imtmetyje ÔĀģ Tuo metu jau buvo Ňĺinoma, kad pagrindiniai lńÖsteliŇ≥ makrokomponentai yra DNR, RNR, baltymai ir angliavandeniai ÔĀģ IŇ° IIIS tipo bakterijŇ≥ jie paruoŇ°ńó lńÖsteliŇ≥ ekstraktus, turinńćius kiekvienńÖ iŇ° Ň°iŇ≥ makromolekuliŇ≥ ÔĀģ Tiktai ekstraktai, turintys iŇ°grynintńÖ DNR, galńójo transformuoti IIR tipo bakterijas ńĮ IIIS tipo bakterijas Avery, MacLeod ir McCarty eksperimentai 2-10
- 11. 2-11 ÔĀģ Avery et al taip pat atliko ir kitńÖ eksperimentńÖ ÔĀģ Jo reikńójo norint ńĮrodyti, kad bŇętent DNR, o ne koks nors kitas ekstrakto komponentas (RNR ar baltymas) yra genetinńó medŇĺiaga
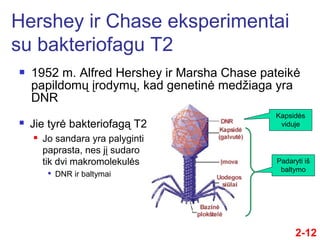
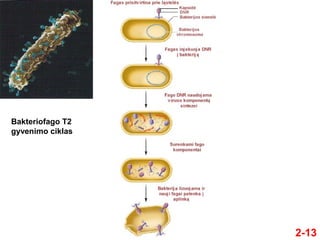
- 12. ÔĀģ 1952 m. Alfred Hershey ir Marsha Chase pateikńó papildomŇ≥ ńĮrodymŇ≥, kad genetinńó medŇĺiaga yra DNR Hershey ir Chase eksperimentai su bakteriofagu T2 2-12 ÔĀģ Jie tyrńó bakteriofagńÖ T2 ÔĀģ Jo sandara yra palyginti paprasta, nes jńĮ sudaro tik dvi makromolekulńós ÔĀģ DNR ir baltymai Padaryti iŇ° baltymo Kapsidńós viduje
- 14. ÔĀģ Hershey and Chase eksperimentŇ≥ santrauka: ÔĀģ Naudojo radioizotopus, norńódami atskirti DNR nuo baltymŇ≥ ÔĀģ 32 P specifiŇ°kai paŇĺymi DNR ÔĀģ 35 S specifiŇ°kai paŇĺymi baltymus ÔĀģ Radioaktyviai paŇĺymńótais fagais buvo uŇĺkreńćĺĪ≤Ļmos neradioaktyvios Escherichia coli lńÖstelńós ÔĀģ Po tam tikro laiko, reikalingo infekcijai ńĮvykti, likusios fagŇ≥ dalelńós buvo paŇ°alinamos ÔĀģ => FagŇ≥ liekanos ir E. coli lńÖstelńós buvo atskiriamos ÔĀģ Radioaktyvumas buvo vertinamas scintiliaciniu skaitikliu 2-14
- 15. Hipotezńó ÔĀģ Tik fago genetinńó medŇĺiaga yra injekuojama ńĮ bakterijas ÔĀģ Radioaktyvi Ňĺymńó turi parodyti, ar tai yra baltymas, ar DNR 2-15
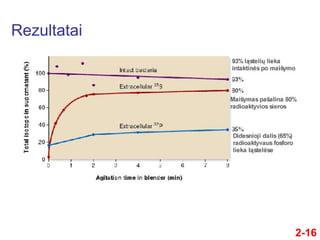
- 16. Rezultatai 2-16
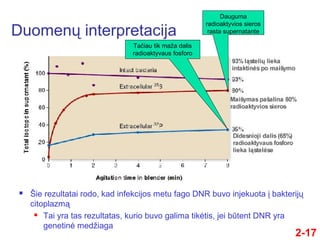
- 17. DuomenŇ≥ interpretacija 2-17 ÔĀģ ҆ie rezultatai rodo, kad infekcijos metu fago DNR buvo injekuota ńĮ bakterijŇ≥ citoplazmńÖ ÔĀģ Tai yra tas rezultatas, kurio buvo galima tikńótis, jei bŇętent DNR yra genetinńó medŇĺiaga Dauguma radioaktyvios sieros rasta supernatante TańćĺĪ≤Ļu tik maŇĺa dalis radioaktyvaus fosforo
- 18. 2-18 ÔĀģ 1956 m. A. Gierer ir G. Schramm iŇ°skyrńó RNR iŇ° augalŇ≥ viruso ‚Äď tabako mozaikos viruso (TMV) ÔĀģ IŇ°valyta RNR sukeldavo tas pańćĺĪ≤Ļs paŇĺaidas, kaip ir intaktinis TMV ÔĀģ Taigi, Ň°iŇ≥ virusŇ≥ genomńÖ sudaro RNR ÔĀģ Dabar aptikta nemaŇĺai RNR virusŇ≥ Kai kuriuose virusuose genetinńós medŇĺiagos funkcijńÖ atlieka RNR
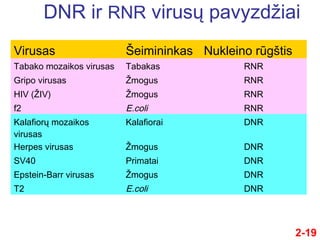
- 19. DNR ir RNR virusŇ≥ pavyzdŇĺiai Virusas ҆eimininkas Nukleino rŇęgŇ°tis Tabako mozaikos virusas Tabakas RNR Gripo virusas ŇĹmogus RNR HIV (ŇĹIV) ŇĹmogus RNR f2 E.coli RNR KalafiorŇ≥ mozaikos virusas Kalafiorai DNR Herpes virusas ŇĹmogus DNR SV40 Primatai DNR Epstein-Barr virusas ŇĹmogus DNR T2 E.coli DNR 2-19
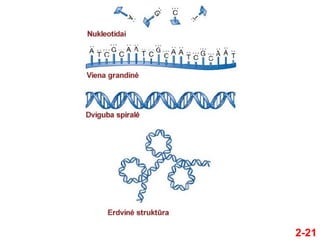
- 20. ÔĀģ DNR ir RNR yra stambios makromolekulńós, susidaranńćios iŇ° keleto lygmenŇ≥ struktŇęrŇ≥ ÔĀģ 1. Nukleotidai sudaro pasikartojanńćius vienetus ÔĀģ 2. Nukleotidai susijungia sudarydami grandinńô ÔĀģ 3. Dvi grandinńós sńÖveikauja sudarydamos dvigubńÖ spiralńô ÔĀģ 4. Dviguba spiralńó susisuka, susilanksto ir sńÖveikauja su baltymais, sudarydama 3-D struktŇęras, formuojanńćĺĪ≤Ļs chromosomas 2.2 NUKLEINO RŇ™G҆ńĆIŇ≤ STRUKTŇ™RA 2-20
- 21. 2-21
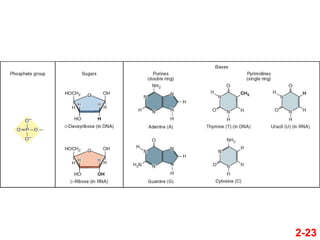
- 22. 2-22 ÔĀģ Nukleotidai yra pasikartojantys DNR ir RNR struktŇęriniai vienetai ÔĀģ Juos sudaro trys komponentai ÔĀģ Fosfatinńó grupńó ÔĀģ Pentozńó (cukrus) ÔĀģ Azotinńó bazńó Nukleotidai
- 23. 2-23
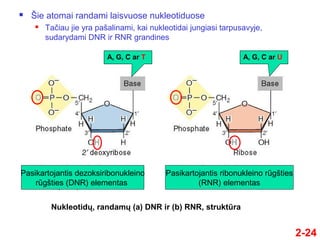
- 24. 2-24 NukleotidŇ≥, randamŇ≥ (a) DNR ir (b) RNR, struktŇęra A, G, C ar T ÔĀģ ҆ie atomai randami laisvuose nukleotiduose ÔĀģ TańćĺĪ≤Ļu jie yra paŇ°alinami, kai nukleotidai jungiasi tarpusavyje, sudarydami DNR ir RNR grandines A, G, C ar U Pasikartojantis dezoksiribonukleino rŇęgŇ°ties (DNR) elementas Pasikartojantis ribonukleino rŇęgŇ°ties (RNR) elementas
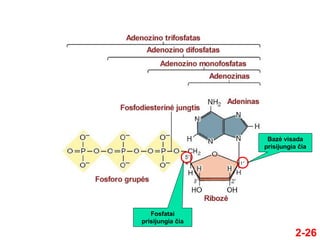
- 25. 2-25 ÔĀģ Bazńó + cukrus ÔɆ nukleozidas ÔĀģ PavyzdŇĺiai ÔĀģ Adeninas + ribozńó = adenozinas ÔĀģ Adeninas + dezoksiribozńó = dezoksiadenozinas ÔĀģ Bazńó + cukrus + fosfatas(i) ÔɆ nukleotidas ÔĀģ PavyzdŇĺiai ÔĀģ Adenozino monofosfatas (AMP) ÔĀģ Adenozino difosfatas (ADP) ÔĀģ Adenozino trifosfatas (ATP)
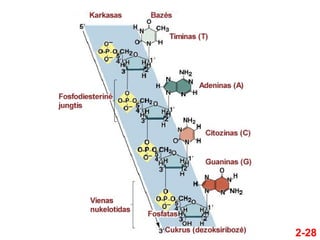
- 27. 2-27 ÔĀģ Nukleotidai kovalentiŇ°kai susijungia per fosfodiesterinńô jungtńĮ ÔĀģ Fosfatas jungia vieno nukleotido 5‚Äô anglies atomńÖ su kito nukleotido 3‚Äô anglies atomu ÔĀģ Tuo bŇędu grandinńó turi kryptńĮ ÔĀģ IŇ° 5‚Äô ńĮ 3‚Äô ÔĀģ Fosforo rŇęgŇ°ties liekanos ir cukrŇęs sudaro nukleino rŇęgŇ°ties grandinńós karkasńÖ ÔĀģ Bazńós kyŇ°o iŇ° karkaso
- 28. 2-28
- 29. 2-29 ÔĀģ 1953 m. James Watson ir Francis Crick nustatńó, kad DNR sudaro dviguba spiralńó, kurioje nukleotidai sńÖveikauja tarpusavyje pagal tam tikras taisykles ÔĀģ TańćĺĪ≤Ļu Ň°io atradimo mokslinius pagrindus sukŇęrńó kiti mokslininkai ÔĀģ Linus Pauling ÔĀģ Rosalind Franklin ir Maurice Wilkins ÔĀģ Erwin Chargaff DNR struktŇęra buvo iŇ°aiŇ°kinta remiantis keliais esminiais rezultatais
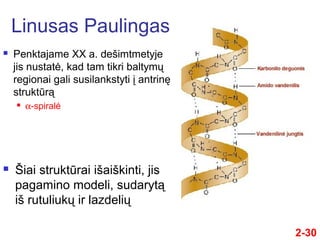
- 30. 2-30 ÔĀģ Penktajame XX a. deŇ°imtmetyje jis nustatńó, kad tam tikri baltymŇ≥ regionai gali susilankstyti ńĮ antrinńô struktŇęrńÖ ÔĀģ őĪ-spiralńó Linusas Paulingas ÔĀģ ҆iai struktŇęrai iŇ°aiŇ°kinti, jis pagamino modeli, sudarytńÖ iŇ° rutuliukŇ≥ ir lazdeliŇ≥
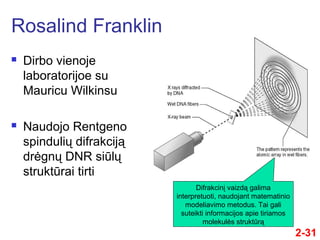
- 31. 2-31 ÔĀģ Dirbo vienoje laboratorijoe su Mauricu Wilkinsu ÔĀģ Naudojo Rentgeno spinduliŇ≥ difrakcijńÖ drńógnŇ≥ DNR siŇęlŇ≥ struktŇęrai tirti Rosalind Franklin DifrakcinńĮ vaizdńÖ galima interpretuoti, naudojant matematinio modeliavimo metodus. Tai gali suteikti informacijos apie tiriamos molekulńós struktŇęrńÖ
- 32. 2-32 ÔĀģ Rentgeno difrakcijos metodais ji pasiekńó gana didelńós paŇĺangos, tyrinńójant DNR struktŇęrńÖ ÔĀģ Difrakciniai vaizdai atspindńójo kai kurias pagrindines DNR savybes ÔĀģ Spiralinńó struktŇęra ÔĀģ Daugiau nei viena grandinńó ÔĀģ 10 baziŇ≥ porŇ≥ tenka vienai pilnai spiralńós vijai Rosalind Franklin
- 33. 2-33 ÔĀģ Chargaffas pirmasis panaudojo daugelńĮ nukleino rŇęgŇ°ńćiŇ≥ iŇ°skyrimo iŇ° gyvŇ≥ lńÖsteliŇ≥, iŇ°gryninimo ir matavimo biocheminiŇ≥ metodŇ≥ ÔĀģ Tuo metu jau buvo Ňĺinoma, kad DNR sudaro keturios bazńós: A, G, C ir T ÔĀģ BaziŇ≥ sudńóties analizńó gali atskleisti svarbias DNR struktŇęros ypatybes Erwino Chargaffo eksperimentai
- 34. 2-34
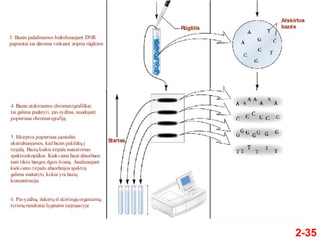
- 35. 2-35
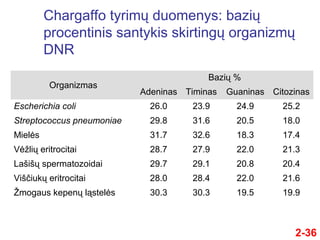
- 36. Chargaffo tyrimŇ≥ duomenys: baziŇ≥ procentinis santykis skirtingŇ≥ organizmŇ≥ DNR Organizmas BaziŇ≥ % Adeninas Timinas Guaninas Citozinas Escherichia coli 26.0 23.9 24.9 25.2 Streptococcus pneumoniae 29.8 31.6 20.5 18.0 Mielńós 31.7 32.6 18.3 17.4 VńóŇĺliŇ≥ eritrocitai 28.7 27.9 22.0 21.3 LaŇ°iŇ°Ň≥ spermatozoidai 29.7 29.1 20.8 20.4 ViŇ°ńćiukŇ≥ eritrocitai 28.0 28.4 22.0 21.6 ŇĹmogaus kepenŇ≥ lńÖstelńós 30.3 30.3 19.5 19.9 2-36
- 37. RezultatŇ≥ interpretavimas 2-37 ÔĀģ Duomenys, pateikti lentelńóje, sudaro tik nedidelńô dalńĮ visŇ≥ rezultatŇ≥, gautŇ≥ Chargaffo ÔĀģ IŇ° rezultatŇ≥ akivaizdŇĺiai matyti, kad ÔĀģ Adenino procentas = timino procentui ÔĀģ Citozino procentas = guanino procentui ÔĀģ ҆is dńósningumas vadinamas Chargaffo taisykle ÔĀģ Tai buvo vienas pagrindiniŇ≥ atspirties taŇ°kŇ≥, kuriuos panaudojo Watsonas ir Crickas DNR struktŇęrai iŇ°aiŇ°kinti
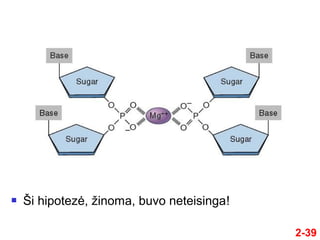
- 38. 2-38 ÔĀģ BŇędami susipaŇĺinńô su visais Ň°iais tyrimŇ≥ rezultatais, Watsonas ir Crickas pasiryŇĺo iŇ°siaiŇ°kinti DNR struktŇęrńÖ ÔĀģ Jie bandńó pagaminti DNR modelńĮ iŇ° rutuliukŇ≥ ir lazdeliŇ≥, kuriam bŇętŇ≥ bŇędingi visi Ňĺinomi eksperimentiniai DNR bruoŇĺai ÔĀģ Esminis klausimas buvo kaip dvi (ar daugiau) grandiniŇ≥ sńÖveikauja tarpusavyje ÔĀģ Pirminńó idńója buvo, kad grandinńós sńÖveikauja per fosfato- Mg++ sńÖsiuvas Watsonas ir Crickas
- 39. 2-39 ÔĀģ ҆i hipotezńó, Ňĺinoma, buvo neteisinga!
- 40. 2-40 ÔĀģ Tada jie sukŇęrńó modelńĮ, kuriame ÔĀģ Fosfodezoksiribozinis karkasas buvo iŇ°orńóje ÔĀģ Bazńós buvo projektuojamos viena prieŇ°ais kitńÖ ÔĀģ PradŇĺioje jie manńó, kad vandenilinńós jungtys jungia identiŇ°kas bazes, esanńćĺĪ≤Ļs prieŇ°ingose grandinńóse ÔĀģ t.y., A su A, T su T, C su C, and G su G ÔĀģ TańćĺĪ≤Ļu modeliavimas parodńó, kad ir Ň°is modelis nńóra teisingas Watsonas ir Crickas
- 41. 2-41 ÔĀģ Tik tada jie suprato, kad vandenilinńós jungtys susidaro tarp A ir T bei tarp C ir G ÔĀģ Taigi, jie pagamino modelńĮ, kuriame A sńÖveikavo su T, o G ‚Äď su C ÔĀģ ҆is modelis jau atitiko visus tuo metu Ňĺinomus duomenis apie DNR struktŇęrńÖ Watsonas ir Crickas
- 42. ÔĀģWatsonas, Crickas ir Wilkinsas 1962 m. uŇĺ DNR struktŇęros iŇ°aiŇ°kinimńÖ gavo Nobelio premijńÖ ÔĀģDeja, Rosalind Franklin mirńó 1958, o Nobelio premijos nńóra skiriamos po mirties. Watsonas ir Crickas 2-42
- 43. 2-43 ÔĀģ Pagrindinńós struktŇęros ypatybńós Dviguba DNR spiralńó ÔĀģ Dvi grandinńós yra susivijusios apie centrinńô aŇ°ńĮ ÔĀģ VienńÖ pilnńÖ apvijńÖ sudaro 10 baziŇ≥ ÔĀģ Abidvi grandinńós yra antiparalelios ÔĀģ Vienos kryptis yra nuo 5‚Äô ńĮ 3‚Äô, o kitos - nuo 3‚Äô ńĮ 5‚Äô ÔĀģ Spiralńó yra deŇ°ninńó ÔĀģ Kai ji vijasi nuo jŇęsŇ≥, spiralńó sukasi pagal laikrodŇĺio rodyklńô
- 44. 2-44 ÔĀģ Pagrindinńós struktŇęros ypatybńós Dviguba DNR spiralńó ÔĀģ DNR struktŇęra yra stabilizuojama ÔĀģ 1. Tarp komplementariŇ≥ baziŇ≥ susidaro vandenilinńós jungtys ÔĀģ A jungiasi su T per dvi vandenilines jungtis ÔĀģ C jungiasi su G per tris vandenilines jungtis ÔĀģ 2. BaziŇ≥ stekingas ÔĀģ DNR struktŇęroje bazńós yra orientuotos taip, kad jŇ≥ plokŇ°ńćiosios dalys ‚ÄúŇĺiŇęri‚ÄĚ viena ńĮ kitńÖ. Esant tokiai orientacijai, tarp baziŇ≥ susidaro stekingo jńógos, kurios papildomai stabilizuoja DNR struktŇęrńÖ
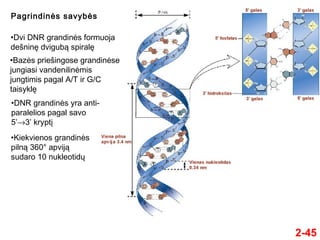
- 45. 2-45 Pagrindinńós savybńós ‚ÄĘDvi DNR grandinńós formuoja deŇ°ninńô dvigubńÖ spiralńô ‚ÄĘBazńós prieŇ°ingose grandinńóse jungiasi vandenilinńómis jungtimis pagal A/T ir G/C taisyklńô ‚ÄĘDNR grandinńós yra anti- paralelios pagal savo 5‚Äô‚Üí3‚Äô kryptńĮ ‚ÄĘKiekvienos grandinńós pilnńÖ 360¬į apvijńÖ sudaro 10 nukleotidŇ≥
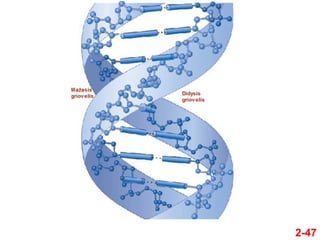
- 46. 2-46 ÔĀģ Pagrindinńós struktŇęros ypatybńós Dviguba DNR spiralńó ÔĀģ Spiralńós pavirŇ°iuje susiformuoja du asimetriŇ°ki grioveliai ÔĀģ 1. Didysis griovelis ÔĀģ 2. MaŇĺasis griovelis ÔĀģ Kai kurie baltymai gali jungtis prie Ň°iŇ≥ grioveliŇ≥ ÔĀģ Tuo bŇędu jie gali sńÖveikauti su tam tikromis nukleotidŇ≥ sekomis
- 47. 2-47
- 48. 2-48 ÔĀģ DNR dviguba spiralńó gali sudaryti skirtingos konformacijos antrines struktŇęras ÔĀģ Dominuojanti forma, randama gyvose lńÖstelńóse yra B-DNR ÔĀģ Esant tan tikroms in vitro sńÖlygoms, gali formuotis A-DNR ir Z-DNR dvigubos spiralńós DNR gali susiformuoti ńĮ keliŇ≥ tipŇ≥ dvigubas spirales
- 49. 2-49 ÔĀģ A-DNR ÔĀģ DeŇ°ninńó spiralńó ÔĀģ Vienoje vijoje 11 bp ÔĀģ Susiformuoja esant Ňĺemai drńógmei ÔĀģ Nńóra aiŇ°kiŇ≥ ńĮrodymŇ≥ apie jos biologinńô svarbńÖ ÔĀģ Z-DNR ÔĀģ Kairinńó spiralńó ÔĀģ Vienoje vijoje 12 bp ÔĀģ FormavimńÖ skatina ÔĀģ Esant didelei druskŇ≥ koncentracijai - GC-turtingos sekos ÔĀģ Esant maŇĺai druskŇ≥ koncentracija ‚Äď citozino metilinimas ÔĀģ Tokia konformacija gali bŇęti svarbi transkripcijai ir rekombinacijai, bent jau mieliŇ≥ lńÖstelńóse
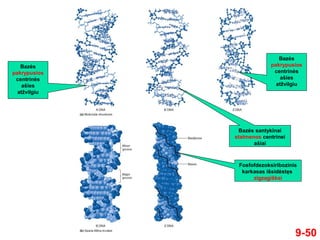
- 50. 9-50 Bazńós pakrypusios centrinńós aŇ°ies atŇĺvilgiu Bazńós pakrypusios centrinńós aŇ°ies atŇĺvilgiu Fosfofdezoksiribozinis karkasas iŇ°sidńóstńôs zigzagiŇ°kai Bazńós santykinai statmenos centrinei aŇ°iai
- 51. 2-51 ÔĀģ XX a. ҆eŇ°tojo deŇ°imtmeńćio pabaigoje Alexander Rich et al atrado trigubńÖ DNR ÔĀģ Ji susiformavo in vitro naudojnant sintetinńós DNR fragmentus ÔĀģ Devintajame deŇ°imtmetyje buvo nustatyta, kad natŇęrali dvigrandininńó DNR gali jungtis su sintetine DNR grandine ir suformuoti trigubńÖ DNR spiralńô (tripleksńÖ) ÔĀģ Sintetinńó DNR grandinńó jungiasi prie natŇęralios DNR didŇĺiojo griovelio DNR gali sudaryti trigubńÖ spiralńô
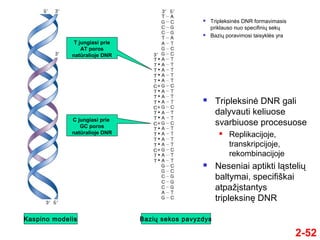
- 52. 2-52 ÔĀģ Tripleksinńós DNR formavimasis priklauso nuo specifiniŇ≥ sekŇ≥ ÔĀģ BaziŇ≥ poravimosi taisyklńós yra ÔĀģ Tripleksinńó DNR gali dalyvauti keliuose svarbiuose procesuose ÔĀģ Replikacijoje, transkripcijoje, rekombinacijoje ÔĀģ Neseniai aptikti lńÖsteliŇ≥ baltymai, specifiŇ°kai atpaŇĺńĮstantys tripleksinńô DNR T jungiasi prie AT poros natŇęralioje DNR C jungiasi prie GC poros natŇęralioje DNR Kaspino modelis BaziŇ≥ sekos pavyzdys
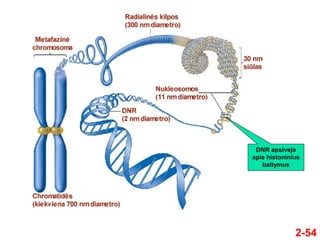
- 53. 2-53 ÔĀģ Tam, kad DNR tilptŇ≥ lńÖstelńójńó, ji turi bŇęti efektyviai kompaktizuota per erdvines struktŇęras ÔĀģ Tokios struktŇęros sukuriamos dalyvaujant baltymams, galintiems jungtis prie DNR ÔĀģ PlańćĺĪ≤Ļu apie tai bus kalbama sekanńćioje paskaitoje Erdvinńó DNR struktŇęra
- 55. 2-55 ÔĀģ Pirminńó RNR grandinńós struktŇęra yra labai panaŇ°i ńĮ DNR grandinńós struktŇęrńÖ ÔĀģ RNR grandinńós paprastai bŇęna nuo keliŇ≥ Ň°imtŇ≥ iki keliŇ≥ tŇękstanńćiŇ≥ nukleotidŇ≥ ilgio ÔĀģ RNR sintetinama naudojant DNR kaip matricńÖ. ҆ios sintezńós metu naudojama tik viena iŇ° dviejŇ≥ DNR grandiniŇ≥ RNR struktŇęra
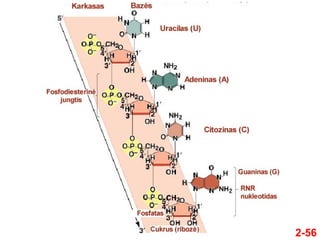
- 56. 2-56
- 57. 2-57 ÔĀģ Nors RNR daŇĺniausiai yra viengrandininńó, RNE molekulńóse gali formuotis dvigrandininiai regionai ÔĀģ ҆ios antrinńós struktŇęros susidaro dńól komplementariŇ≥ baziŇ≥ poravimosi ÔĀģ A su U ir C su G ÔĀģ Dńól to trumpuose RNR segmentuose susiformuoja dviguba spiralńó ÔĀģ TipiŇ°ka RNR dviguba spiralńó ÔĀģ Yra deŇ°ninńó ÔĀģ Sudaro A formńÖ, kurioje vienai vijai tenka 11-12 bp ÔĀģ Gali susidaryti skirtingŇ≥ tipŇ≥ RNR antrinńós struktŇęros
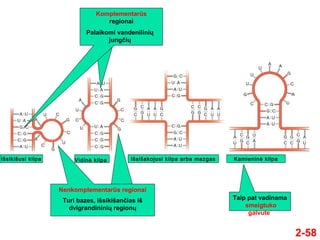
- 58. 2-58 Taip pat vadinama smeigtuko galvute KomplementarŇęs regionai NenkomplementarŇęs regionai Palaikomi vandeniliniŇ≥ jungńćiŇ≥ Turi bazes, iŇ°sikiŇ°anńćĺĪ≤Ļs iŇ° dvigrandininiŇ≥ regionŇ≥ IŇ°sikiŇ°usi kilpa Vidinńó kilpa IŇ°siŇ°akojusi kilpa arba mazgas Kamieninńó kilpa
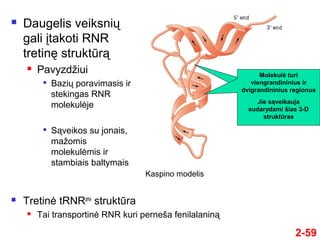
- 59. 2-59 ÔĀģ Daugelis veiksniŇ≥ gali ńĮtakoti RNR tretinńô struktŇęrńÖ ÔĀģ PavyzdŇĺiui ÔĀģ BaziŇ≥ poravimasis ir stekingas RNR molekulńóje ÔĀģ SńÖveikos su jonais, maŇĺomis molekulńómis ir stambiais baltymais ÔĀģ Tretinńó tRNRphe struktŇęra ÔĀģ Tai transportinńó RNR kuri perneŇ°a fenilalaninńÖ Molekulńó turi viengrandininius ir dvigrandininius regionus Jie sńÖveikauja sudarydami Ň°ias 3-D struktŇęras Kaspino modelis